Carnivorous Plants
_______
I. Anatomy of Carnivory
2017-03-15
Updated 2020-11-12
FOR CARNIVORY TO TAKE EFFECT, there must be contact between leaf and prey. If a
fly alights on a leaf and a moment later flies away, the contact is not
sufficient. A fly must be persuaded to stay for a period during which the
digestion can take place.
The first step in the sequence of events leading to
successful carnivory is a lure
that brings prey to physical proximity.
Capture
follows (forced detention).
Digestion then starts
that decomposes the prey. The digestion proceeds with
plant's own enzyme or with the help of other commensal organisms.
Finally, absorption
takes place that carries the
products of digestion into the leaf
and to the growth site of the plant.
(1)
Lure --> (2)
Capture --> (3)
Digestion --> (4)
Absorption
The main nutrients selectively
absorbed by carnivorous plants are nitrogen (N) and phosphorus (P)
-- major plant nutrients -- but some
other elements required in trace amounts such as potassium (K:
kalium) and
magnesium (Mg) are also utilized by some species.
Trap Mechanism
There are four major trap types used by carnivorous plants: Pitfall trap,
adhesive trap, snap-trap, and suction trap (with another somewhat related
lobster-pot trap). We tend to emphasize the trap mechanisms when we
analyze relatedness of different carnivorous plants, but digestion/absorption schemes
that they use are just as
significant in their evolutionary pathway.
Glands A
gland is a structure which secretes fluids. Normally
the walls of the
epidermal cells are covered with a cuticle to reduce water loss, but the outer cells of
a gland have a thin, perforated cuticle.
Glands play a significant role in carnivory: Glands are used for lure (nectaries
- nectar
glands), digestion (enzyme secretion) and absorption of the digestion products. For
adhesive traps, the
glands are also used to
secrete viscous mucilage (or resin in the case of Roridula).
Depending on the types of traps, different kinds of glands perform different
functions, or the same gland assumes multiple tasks.
Digestion In
pitfall traps, the digestion can start in the preformulated fluids immediately
upon prey capture. In other trap types, there is detection/perception of stimuli
associated with prey capture, and the digestive process is initiated within a
certain time frame. The absorption of the products of digestion is usually
carried out by the same digestive glands.
In the case
of some traps that do not possess any glands for absorption, cuticular discontinuities provide passage of
nutrients into the leaf tissue.
Repeatability Generally, adhesive secretion repeats many times, but may occur only once, as in Pinguicula’s adhesive glands. Likewise, the secretion of digestive enzyme may repeat several times, as in Dionaea, or can occur only once, as in Pinguicula, depending on the gland anatomy (presence or absence of conducting tissues to the gland).
EVOLUTION
CARNIVOROUS PLANTS (CP) EMERGED FROM non-carnivores at several distinct points in evolution. They only
appeared as either a pitfall or adhesive trap -- and then, only
some adhesives moved on to
more sophisticated trap mechanisms.
CP emerged as pitfalls in:
----- In the
order Ericales:
Darlingtonia / Sarracenia / Heliamphora
----- In the order
Oxalidales:
Cephalotus
CP emerged as adhesive traps in:
-----
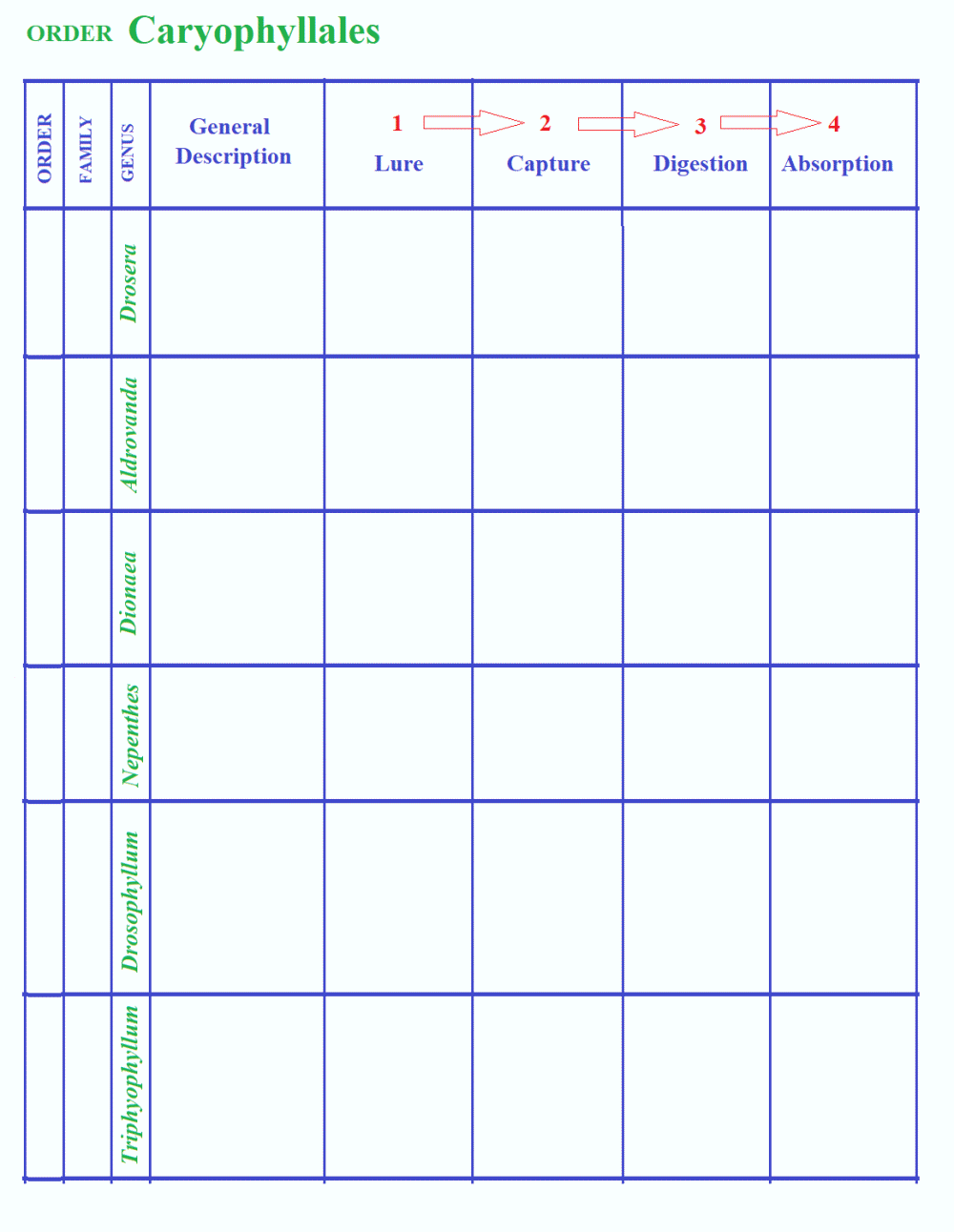
In the order Caryophyllales:
Drosera
>> Snap-traps (Aldrovanda / Dionaea)
>> Pitfall (Nepenthes) >>
Adhesive again (Drosophyllum >>Triphyophyllum)
----- In the order
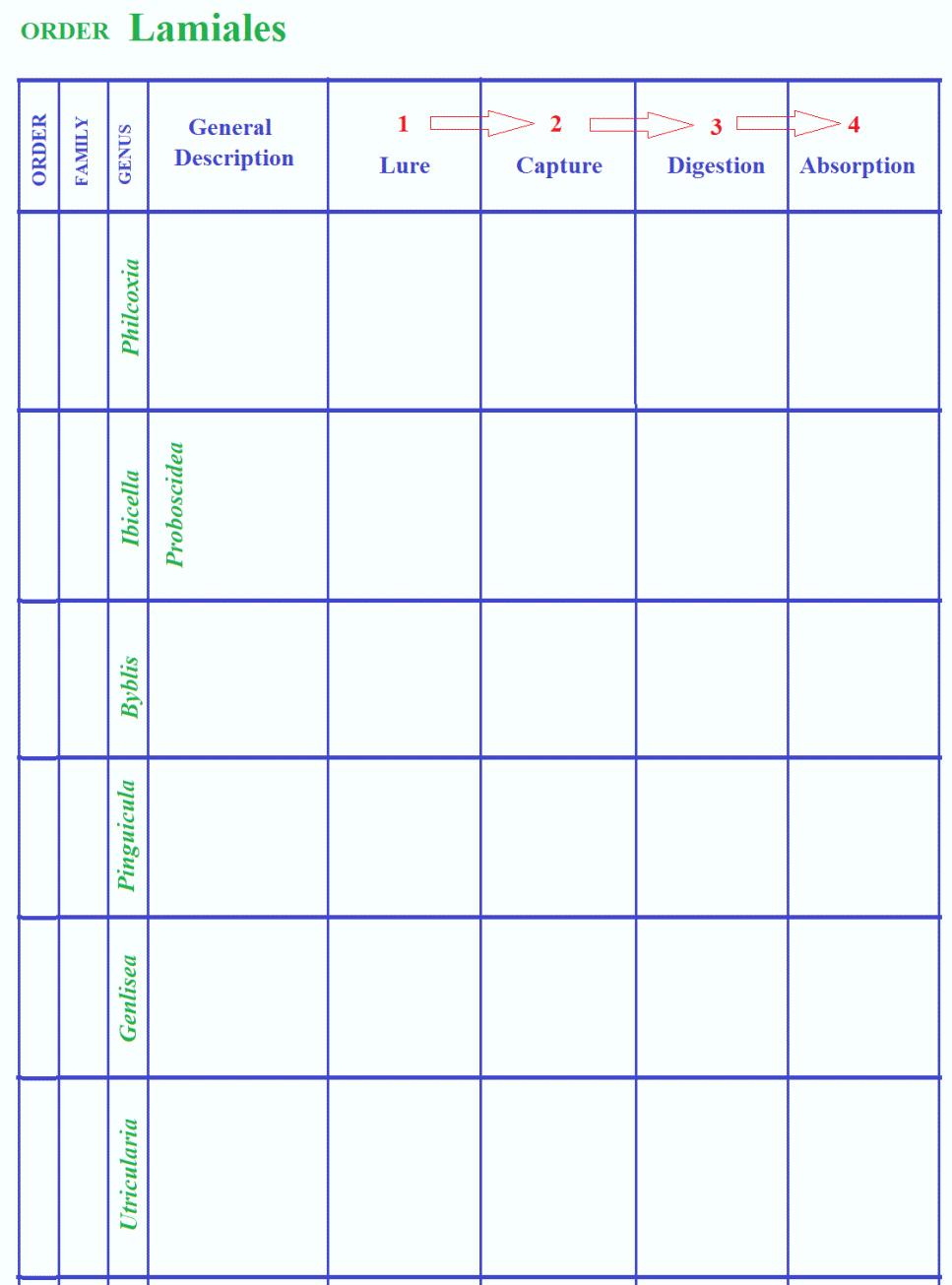
Lamiales:
Pinguicula
>> Lobster pot (Genlisea) >>
Suction trap (Utricularia)
Ibicella /
Proboscidea --- x
Byblis ---
x
Philcoxia
--- x
----- In the order
Ericales:
Roridula
--- x
(* Molecular systematics places Roridula
and Sarraceniaceae in the same Ericales order and they are closely related.
While Roridula is adhesive (resin) and Sarracenia/Darlingtonia/Heliamphora
are pitfall, both families share a very
similar absorption mechanism - the efficient absorption of the prey-derived
nutrients is achieved by the cuticular
discontinuity of the leaf surface.)
Aquatic Environment Pushed
Evolution
In sundews (order Caryophyllales) and butterworts (order Lamiales),
both of which are adhesive carnivores, the transition to aquatic environment seems
to have exerted pressure to move forward to more sophisticated trapping schemes
(provided we assume Drosera-Aldrovanda-Dionaea evolution order).
Movement in Capture
We
note that in the course of adaptation to carnivory, there are two distinct points at which physical
movement was acquired during prey capture, leading to so-called "active" traps in carnivorous
plants. One is in the order Caryophyllales in the branch leading to Drosera --
namely the Drosera's tentacle bending (and leaf folding) that eventually
led to the snap-traps of Aldrovanda and Dionaea. In the same order, the
branch leading to Nepenthes, Drosophyllum and Triphyophyllum
either lost or did not acquire any such movement. The other is the leaf movement (curling and
dishing) in Pinguicula upon prey capture that --- though not as direct or
obvious a transition as in Droseraceae --- may have some connection to the
workings of some aspect of Utricularia traps...
Some comments on use of the term "active" : The
active trap simply means some movement is visible to the naked eye. If water
molecules move in the trap, it is not visible. So, a bladderwort trap may be
considered passive -- until it blasts as a result of gradual expulsion of water.
The
distinction between "active" and "passive" traps may be somewhat artificial...
REF: PLANT MOVEMENT IN RESPONSE TO STIMULI
1) Nastic Movement - movement in a pre-determined direction, regardless of the location of stimuli.
2) Tropistic Movement - movement in a direction that has correlation to the direction/location of stimuli.
Phototropism - movement toward light
Gravitropism -
movement in response to gravity
Thigmotropism - movement in response to physical touch
Occurrences of Sticky Carnivores
We often talk about the number of
occurrences of carnivory in the course of angiosperm (flowering plant)
evolution. In the order Lamiales, there appears to be multiple,
independent points at which adhesive-trap carnivores emerged (Philcoxia, Byblis,
Ibicella/Proboscidea, Pinguicula). But there are many non-carnivorous taxa in this order that possess glandular foliage, generally for defense
purpose. These adhesive glands probably can be traced back to a very ancient
common origin. Given what we do not know, once a reasonably efficient and
workable gland structure has been established, creating a sticky carnivore out
of a sticky defense seems a relatively small step. Therefore, even if this final
step toward carnivory might have been undertaken independently in a handful of
different families in Lamiales, considering these occurrences completely
independent is a little misleading (or of little value). We should be cognizant
of the fact that carnivory requires a lot of things and the ground work had started
long ago, probably in the common ancestors of these families. This certainly
does not suggest to imply that "carnivory" had been contemplated upon as a
long-term goal when the gland structures were being developed eons ago and, from
that point forward, having been
worked on to finally come to fruition by this final step --- Evolution, by
definition, does not have any "future" plan. What I am saying is that, once
we understand what carnivory entails and how it might have come about, counting the
number of occurrences of carnivory --- by just noting the final stage toward carnivory
--- may not be as meaningful an endeavor as we might have initially thought.
“Sticky"
Difference
There are many adhesive-trap carnivores that use sticky mucus to capture prey.
They are found in three orders: Caryophyllales, Ericales and Lamiales. It is interesting to note that all adhesive glands of carnivorous plants are
stalked glands raised above the leaf surface by some length; none of them are
sessile glands (stalkless).
Darwin noted some difference between Drosera and Drosophyllum in their mucilage characteristics. In order to highlight this subtle but crucial difference, let us divide sticky-leaf carnivores into two groups. Drosera is the only member in Group 1, and the rest goes to Group 2. This grouping has to do with the way the digestive process is carried out after prey are captured by the adhesive glands.
DUAL-ABILITY OF STALKED GLANDS
Among all sticky-leaf glands, Drosera’s stalked glands (tentacles) are the only glands that have acquired the "dual" ability to perform digestive process (secreting digestive enzymes and absorbing the products of digestion) in addition to prey capture. In all the others, digestion is carried out by separate sessile glands scattered on the leaf surface. This means, for all these carnivorous plants in Group 2, keeping the trapped prey high on the tip of the stalked glands is meaningless, for no digestion will take place there --- the prey must be dropped down on the leaf surface. As noted by some observers (including Darwin), the Drosophyllum’s glue tends to peel off from the gland upon prey capture, exactly for this reason. This is in contrast to the Drosera’s glue that typically remains on the tentacle head to hold the prey during and after prey capture.
By and large, similar trapping behavior can be observed in all adhesive traps belonging to Group 2. In Pinguicula, for instance, the stalked glands readily collapse upon prey capture. This will allow the prey to sink down to the leaf surface and touch the sessile glands. That said, physical contact between prey and digestive glands may not be necessary for digestive process, so long as the small gap between them is bridged with fluids: For digestion, secreted fluids rich with enzyme must touch the body of the prey; for absorption, the fluids containing the digestion products must reach the absorptive glands. Keeping prey on the tall stalked glands of Drosophyllum is not likely to achieve this.
STALKED GLAND MORPHOLOGY
As mentioned above, the mucilage tends to peel off from the gland upon prey
capture in Group 2 carnivores. There may be some difference in mucilage chemical
composition that explains this behavior. Comparing the shape of glands in Group
1 & 2, the gland morphology also seems to support the difference in this behavior...
Group 1 (Drosera) --- The entire, spherical surface of the gland secretes sticky mucilage (i.e., the entire surface has cuticular gaps). As a result of this, the mucilage covers the entire gland surface in sundew tentacles. The contact area between mucilage and gland tends to be large, and therefore the mucilage is more securely attached to the gland.
Group 2 (Drosophyllum, Triphyophyllum, and Pinguicula) --- The gland itself at the stalk tip is flat-shaped, and only the upper surface of the gland secretes mucilage. As such, the mucilage tends to sit on a dish-shaped gland, rather than covering the entire gland. This small contact area may account for the easier detachability of the mucilage from the gland upon prey capture in Drosophyllum.


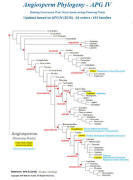
CLASSIFICATION OF CARNIVOROUS PLANTS / CP PHYLOGENY
CARNIVORY AT A GLANCE
(illustration) ------------ under construction
FOR VARIOUS CP GENERA
BELOW, we present the
anatomy of carnivory in four steps showing the basic mechanisms involved.
(1) LURE
- visual (leaf color /
leaf shape / dewy-look... UV absorption pattern)
- olfactory (fragrance)
- food (nectar)
(2) CAPTURE
- pitfall trap (pitcher-shaped leaf
/ waxy surface / slippery hairs / retentive hairs...)
- adhesive trap (adhesive glands - how many
times)
- snap-trap (aquatic
waterwheel plant / terrestrial Venus flytrap)
- lobster-pot trap (glands – water flow
/ hairs)
- suction trap (glands – water expulsion)
(3) DIGESTION
- enzyme secretion by
digestive glands
- the Golgi apparatus (stimuli
detection / how many times...)
- commensals (bacteria
/ commensal organisms / inquilines)
(4) ABSORPTION
- absorption by (digestive) glands.
--- Sessile glands / stoked glands (Drosera only)
- if no glands exist, cuticular discontinuity of epidermis allows absorption
Poales
Bromeliaceae Brocchinia
Catopsis
Eriocaulaceae Paepalanthus
Oxalidales Cephalotaceae Cephlotus
Caryophyllales Droseraceae Drosera
Aldrovanda
Dionaea
Nepenthaceae Nepenthes
Drosophyllaceae Drosophyllum
Dioncophyllaceae Triphyophyllum
Ericales Roridulaceae Roridula
Sarraceniaceae Darlingtonia
Heliamphora
Sarracenia
Lamiales Plantaginaceae Philcoxia
Linderniaceae Lindernia
Byblidaceae Byblis
Martyniaceae Ibicella
Proboscidea
Lentibulariaceae Pinguicula
Genlisea
Utricularia
Copyright (c) 2017 Makoto Honda. All Rights Reserved.