| Update Info Site Index Licensing Images Copyright Notice Privacy Policy Contact Us | |||
|
Introduction
|
Phylogeny of Carnivorous Plants
Phylogenetic systematics attempts to construct a classification of organisms that reflects descent. Molecular genetic research in the past 10 years has greatly enhanced our understanding of evolutionary relationships among angiosperms. Molecular phylogenetic analyses continue to provide quantitative data for objective inference of angiosperm phylogeny, largely eliminating room for subjective interpretations that have resulted in differing taxonomic views in the past.
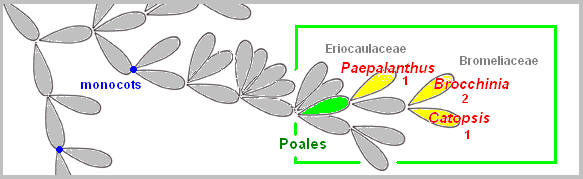
Poales Rather isolated from the rest, the order
Poales (belonging to monocots)
contains carnivorous plants possessing a primitive pitfall trap (Paepalenthus,
Brocchinia, and Catopsis).
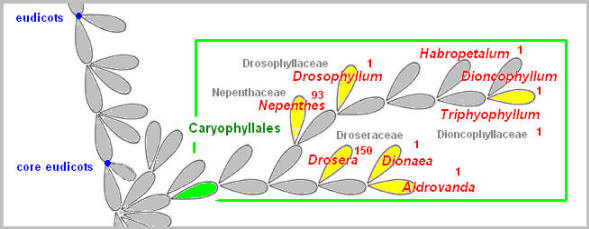
Phylogenetic analyses of DNA sequences have placed Droseraceae (Drosera, Dionaea, and Aldrovanda), Nepenthaceae (Nepenthes), Drosophyllaceae (Drosophyllum) and Dioncophyllaceae (Triphyophyllum) in the same order Caryophyllales. A traditionally accepted close relationship among Drosera, Dionaea, and Aldrovanda based primarily on floral and pollen morphology has been favorably supported in molecular systematics. Sundews (Drosera) cover their leaves with sticky, mucilage-tipped hairs. Their hairs, often referred to as tentacles, are not a simple, single-celled hair. They not only have a multi-cellular stalk, but also possess epidermis as well as vascular systems in the stalk center, as if tentacles themselves are a miniature leaf. Dionaea and Aldrovanda both possess a snap-trap, the most advanced prey trapping mechanism to be found among carnivorous plants. Although Dionaea is a land plant while Aldrovanda is aquatic, they share a great deal of common features in their snap-trap mechanism. The traps have a terminal leaf blade that is divided into two half-shell shaped lobes connected along the midrib. Both traps have sensitive trigger hairs on the inner lobe surface that initiate a swift trap closure when stimulated. Molecular evidence shows these species are closely related, with Aldrovanda being sister to Dionaea, suggesting a common ancestral origin of these snap-traps. Furthermore, Dionaea and Aldrovanda form a clade which is sister to Drosera. This strongly supports a speculation that Dionaea and Aldrovanda evolved from a common sundew-like ancestor. Without any fossil records revealing a hint of intermediate forms, how an ancient adhesive trap developed into a snap-trap remains unknown.
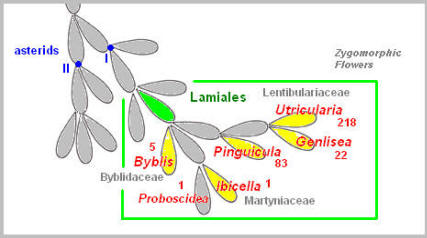
Mutation is likely to have played a role in the development of the snap-trap. Nepenthes contains close to 100 species of tropical pitcher plants from the Old World. The plants develop a pitcher at the tip of a long tendril extending from the midrib of a regular shaped leaf (morphologically, the leaf base). The ancestor of Nepenthes is speculated to be a sundew type plant. The branch leading to Nepenthes has further developed into Drosophyllum which produces sticky glandular leaves, like Drosera. The carnivorous feature then seems to disappear and stay dormant for a while, only to re-emerge in Dioncophyllaceae. The monotypic genus Triphyophyllum is a tropical liana, somewhat reminiscent of Nepenthes, but with slender, adhesive glandular leaves, quite similar to those of Drosophyllum. In Triphyophyllum, these carnivorous leaves are produced only seasonally, and only in the young plant before it enters the adult phase of a climbing liana. The adhesive tentacles of Drosera, Drosophyllum, and Triphyophyllum are all multi-cellular in structure, possess the xylem in the stalk center, and are far more complex than a typical hair commonly found in plants. It is as though, with the acquisition of an adhesive trap trait in the common ancestor of Drosera and Nepenthes, the order Caryophyllales has been predisposed to carnivory. Lamiales Molecular genetics research suggests that in the order Lamiales the transition to carnivory had taken place independently in the three families, Lentibulariaceae, Martyniaceae, and Byblidaceae. Lentibulariaceae contains three genera, each having a morphologically distinct trap type. Two of these, Utricularia and Genlisea, are closely related, both possessing a unique and complex underwater trap to capture tiny water animals (invertebrates).
The other genus Pinguicula has developed an adhesive trap. The surface of the butterwort’s leaf is covered with numerous hairs tipped with mucilage secretions. These hairs are a simple, single-celled hair supporting multi-cellular glands at the tip that produce adhesive mucilage to trap small prey. Martyniaceae (Ibicella) and Byblidaceae (Byblis) have both developed the adhesive trap similar to that of Pinguicula. The hairs of Pinguicula, Byblis and Ibicella (having a multi-cellular stalk) are a simple hair, commonly found in many other plants, and do not possess the complexity of sundew tentacles. The carnivorous plants in this order produce a zygomorphic flower (bilaterally symmetric) though in Byblis it is only partially so. Ericales DNA sequence
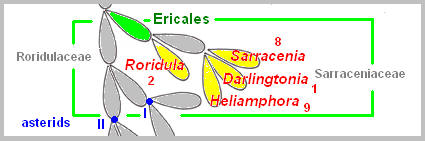
comparisons have placed Sarraceniaceae and Roridulaceae in the order
Ericales. The New World pitcher plant family Sarraceniaceae contains three
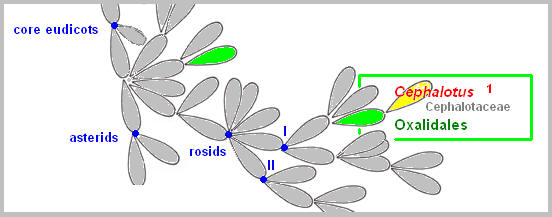
genera, Sarracenia, Darlingtonia, and Heliamphora. Roridula has developed sticky leaves with powerful glue of resin. The plant resembles in shape some species of Drosera, but the similarity is more apparent than real. Roridula’s adhesive trap differs from other flypaper traps in that the secretions is resinous, distinct from water-based, mucilaginous adhesive traps found in other carnivorous species. Oxalidales Cephalotus, an Albany pitcher plant from Western Australia, is placed in the order Oxalidales. The Cephalotus produces two kinds of leaves, a normal, non-carnivorous leaf and a pitcher-shaped pitfall trap. It is seen that the pitcher-shaped leaves of Cephalotus have evolved independently and are not related to other pitfall traps (like Sarracenia and Nepenthes). The similarity is the result of convergent evolution (acquisition of characters resembling each other in function due to similar environmental conditions, not due to common ancestry).
Reference: Hasebe (2005), Angiosperm
Phylogeny Group (2003), Hilu et al.
(2003), and others. |